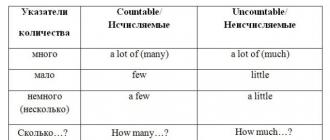
Metode citologice. Rezumat: Metode de cercetare în citologie Următoarele metode nu sunt utilizate în citologie
Principalele metode de studii citologice sunt microscopia luminoasă și electronică, adică utilizarea microscoapelor ușoare și electronice pentru a vedea structurile externe și interne ale celulelor.
Microscoapele ușoare fac, de asemenea, posibilă observarea celulelor vii (de obicei, organisme unicelulare, celule sanguine, sunt folosite pentru aceasta). Cu toate acestea, rezoluția microscoapelor ușoare nu este la fel de mare ca cea a microscoapelor electronice. Rezoluția unui dispozitiv de mărire este distanța minimă dintre două puncte vizibile separat. Pentru microscoapele ușoare această distanță se măsoară în sute de nanometri, iar pentru microscoapele electronice se măsoară în zeci și unități de nanometri. Dacă primul folosește un flux luminos (rezoluția este invers proporțională cu lungimea de undă), atunci cel din urmă folosește un flux de electroni.
Există două tipuri de microscoape electronice - cu transmisie și cu scanare. Rezoluția primei este ceva mai mare, dar cu ajutorul celui de-al doilea puteți obține o imagine tridimensională. Pentru microscoapele cu transmisie se pregătesc secțiuni foarte subțiri prin care trece un fascicul de electroni. În microscoapele cu scanare, un fascicul de electroni este reflectat de un obiect.
Folosit și în studiile citologice metoda microscopiei cu fluorescență, care constă în adăugarea anumitor coloranți la celulele vii, care, atunci când sunt combinați cu diverse componente ale celulei, încep să strălucească. Astfel, este posibilă observarea structurilor celulare (cloroplaste, microtubuli etc.) cu ajutorul unui microscop cu lumină.
Pe lângă microscopie, citologia modernă folosește și alte metode de cercetare. Metoda citochimică vă permite să studiați compoziția chimică a celulelor. Această metodă se bazează pe reacțiile chimice ale anumitor substanțe. Prin adăugarea de reactivi la celule, este posibilă detectarea prezenței ADN-ului, a anumitor proteine etc. în ele, precum și determinarea cantității acestora.
Metoda autoradiografiei presupune introducerea unei substanțe care conține atomi marcați (radioactivi). După ceva timp, moleculele marcate sunt încorporate în biopolimerii celulei și pot fi folosite pentru a urmări progresul proceselor metabolice din celulă.
Începând cu anii 20 ai secolului XX, o astfel de metodă de cercetare citologică ca centrifugare (sau metoda de fracționare a structurilor celulare). Se bazează pe faptul că structurile celulare au mase diferite și, în timpul centrifugării, sedimentează la viteze diferite. Astfel, dacă celulele sunt distruse, atunci după centrifugare amestecul va fi împărțit în fracții, cu structuri mai grele în partea de jos (de obicei nuclee celulare) și mai ușoare în partea de sus.
Relativ nou este metoda culturii celulare, care permite în afara corpului, în condiții special create, să crească celule (colonii) identice din una sau mai multe originale. Această metodă vă permite să studiați proprietățile celulelor sale separat de organism, să efectuați studii citologice, genetice și alte studii.
O nouă metodă de cercetare citologică este metoda microchirurgiei. Folosind un micromanipulator conectat la un microscop, diferite componente sunt îndepărtate sau adăugate din celule și sunt introduse substanțe.
Nivelul celular al organizării vieții
§ 16. Istoria studiului celulelor. Metode de cercetare citologică.
Istoria studiului celulelor.
Lumea celulelor a rămas complet necunoscută până la mijlocul secolului al XVII-lea, când oamenii au învățat să șlefuiască lentilele și să le folosească pentru a îmbunătăți vederea.
Unul dintre primii creatori ai microscopului a fost Robert Hooke fizician, meteorolog, biolog, inginer, arhitect. ÎN 1665 a publicat un album de desene numit „macrografie”, care prezenta observațiile sale la microscop.
Unul dintre contemporanii talentați ai lui Hooke a fost olandezul Anthony van Leeuwenhoek, care a creat 200 de microscoape cu design special propriu. Leeuwenhoek a obținut o creștere de 270 de ori a numărului de obiecte și a făcut descoperiri remarcabile.
Robert Brown în 1833 a descoperit nucleul unei celule. După 1825 Jan Purkinje a dezvoltat metode eficiente de preparare și colorare a preparatelor pentru echipamente microscopice.
Teoria celulară propusă pentru plante în 1837 botanist german Matthias Schleiden,și a fost extins în lumea animală de către prietenul său, fiziologul Theodor Schwann. Puțin mai târziu a fost completat Rudolf Virchow, in care 1885 a formulat propoziția „Fiecare celulă provine dintr-o celulă”.
La mijlocul secolului al XIX-lea. teoria celulară a devenit general acceptată și baza pentru știința celulară - citologie. Până la sfârșitul secolului al XIX-lea. au fost descoperite multe componente ale celulelor. Oamenii de știință le-au descris și le-au dat nume.
Dar în 1945 Citologii au analizat celulele pentru prima dată folosind un microscop electronic și au văzut multe structuri necunoscute anterior. Deci, rolul decisiv în dezvoltarea citologiei revine noilor descoperiri din alte științe, în special din fizică.
Metode de cercetare citologică.
Metoda principală este metoda microscopiei luminoase. Implică utilizarea unui microscop cu lumină, dar numai preparatele citologice special preparate pot fi examinate la microscop cu lumină.
Pentru pregătirea preparatelor, citologii folosesc lame de sticlă și obiecte special pregătite care pot fi examinate.
Cel mai adesea, aceste structuri sunt incolore, așa că trebuie vopsite cu coloranți speciali, diferiți de fiecare dată, în funcție de ce structuri doriți să vedeți.
Există două metode: o metodă de preparare a preparatelor sub presiune - obiectul studiat este pur și simplu zdrobit într-un singur strat între o lamă și o sticlă de acoperire și o metodă de preparare a secțiunilor subțiri constând dintr-un singur strat de celule.
Folosit pentru a studia celulele vii metoda microscopiei cu contrast de fază. Se bazează pe faptul că secțiunile individuale ale unei celule transparente diferă unele de altele în densitate și refracție a luminii.
Când studiază celulele vii, ele folosesc și metoda microscopiei cu fluorescență. Semnificația sa constă în faptul că o serie de substanțe au capacitatea de a străluci atunci când absorb energia luminoasă. De exemplu, dacă examinați celulele plantei printr-un microscop fluorescent, atunci pe corpul albastru închis veți vedea boabe roșii care strălucesc puternic - acestea sunt cloroplaste.
Există o metodă care utilizează izotopi etichetați - metoda autoradiografiei- înregistrarea substanţelor etichetate cu izotopi. Folosind această metodă, puteți vedea ce părți ale celulei primesc substanțe marcate cu izotopi radioactivi.
Metoda microscopiei electronice Citologul a descoperit acele structuri celulare care au dimensiuni mai mici decât lungimea de undă a luminii. Datorită acestei metode, a devenit posibilă examinarea virușilor și organitelor pe care are loc sinteza proteinelor (ribozomi).
De asemenea, citologii pot obține și studia diverse componente ale celulelor folosind fracţionarea celulelor. Celula este mai întâi distrusă, iar apoi structurile celulare sunt izolate folosind un dispozitiv special - o centrifugă.
Metoda de utilizare a culturii celulare este o metodă de depozitare și cultivare pe termen lung în medii nutritive speciale a celulelor, țesuturilor, organelor mici sau părților acestora izolate de corpul uman, animal sau vegetal. Un avantaj important al acestei metode este capacitatea de a observa activitatea vitală a celulelor folosind un microscop.
Importanța metodelor citologice în diagnosticul și tratamentul bolilor umane.
1) Metode citologice sunt folosite în medicină pentru a studia starea fiziologică a corpului uman pe baza studierii structurii celulelor. Sunt folosite pentru a identifica bolile de sânge, recunoașterea tumorilor maligne și benigne, a multor boli ale sistemului respirator, digestiei, urinare, sistemului nervos și tratarea acestora.
2) Celula stem este o celulă imatură capabilă de auto-reînnoire și dezvoltare în celule specializate ale corpului. În corpul adult, celulele stem se găsesc în principal în măduva osoasă și în cantități foarte mici în toate organele și țesuturile. Ele pot fi folosite pentru a trata multe boli.
§ 17. Structura celulelor procariote şi eucariote.
Unitatea structurii celulare.
Conținutul oricărei celule este separat de mediul extern printr-o structură specială - membrana plasmatica (plasmalema). Această izolare vă permite să creați un mediu cu totul special în interiorul celulei, spre deosebire de ceea ce o înconjoară. Prin urmare, procesele care nu au loc în altă parte pot avea loc în celulă; ele sunt numite procesele vieții.
Mediul intern al unei celule vii, delimitat de membrana plasmatică, se numește citoplasmă. Include hialoplasma(substanță transparentă de bază) și organele celulare, precum și diverse structuri nepermanente - incluziuni. Organelele care sunt prezente în orice celulă includ, de asemenea ribozomi, unde se întâmplă sinteza proteinei.
Structura celulelor eucariote.
eucariote- Acestea sunt organisme ale căror celule au un nucleu. Miez- acesta este chiar organelul celulei eucariote în care se stochează informația ereditară înregistrată în cromozomi și din care se transcrie informațiile ereditare. Cromozom este o moleculă de ADN integrată cu proteine. Miezul contine nucleol- locul unde se formează alte organite importante implicate în sinteza proteinelor - ribozomi. Dar ribozomii se formează doar în nucleu și funcționează (adică sintetizează proteine) în citoplasmă. Unele dintre ele sunt libere în citoplasmă, iar altele sunt atașate de membrane, formând o rețea, care se numește endoplasmatic.
Ribozomi- organele nemembranare.
Reticulul endoplasmatic este o rețea de tubuli mărginiți de membrană. Există două tipuri: netede și granulare. Ribozomii sunt localizați pe membranele reticulului endoplasmatic granular, astfel încât proteinele sunt sintetizate și transportate acolo. Și reticulul endoplasmatic neted este locul sintezei și transportului carbohidraților și lipidelor. Nu există ribozomi pe el.
Sinteza proteinelor, carbohidraților și grăsimilor necesită energie, care este produsă în celula eucariotă de „stațiile energetice” ale celulei - mitocondriile.
Mitocondriile- organele cu membrană dublă în care are loc procesul de respirație celulară. Compușii organici sunt oxidați pe membranele mitocondriale și energia chimică se acumulează sub formă de molecule speciale de energie (ATP).
Există, de asemenea, un loc în celulă unde se pot acumula compuși organici și de unde pot fi transportați - acesta este Aparate Golgi, sistem de pungi cu membrană plate. Este implicat în transportul proteinelor, lipidelor și carbohidraților. Aparatul Golgi produce, de asemenea, organele pentru digestia intracelulară - lizozomi.
Lizozomi- organele cu o singură membrană, caracteristice celulelor animale, conțin enzime care pot descompune proteinele, carbohidrații, acizii nucleici și lipidele.
O celulă poate conține organele care nu au o structură de membrană, cum ar fi ribozomi și un citoschelet.
Citoscheletul- acesta este sistemul musculo-scheletic al celulei, include microfilamente, cili, flageli, centrul celular, care produce microtubuli si centrioli.
Există organite caracteristice doar celulelor vegetale - plastide. Există: cloroplaste, cromoplaste și leucoplaste. Procesul de fotosinteză are loc în cloroplaste.
De asemenea, în celulele vegetale vacuole- produse reziduale ale celulei, care sunt rezervoare de apă și compuși dizolvați în ea. Organismele eucariote includ plante, animale și ciuperci.
Structura celulelor procariote.
procariote- organisme unicelulare ale căror celule nu au nucleu.
Celulele procariote sunt de dimensiuni mici și stochează material genetic sub forma unei molecule circulare de ADN (nucleoid). Organismele procariote includ bacterii și cianobacteriile, care anterior erau numite alge albastre-verzi.
Dacă procesul de respirație aerobă are loc la procariote, atunci se folosesc proeminențe speciale ale membranei plasmatice pentru aceasta - mezosomi. Dacă bacteriile sunt fotosintetice, atunci procesul de fotosinteză are loc pe membranele fotosintetice - tilacoizi.
Sinteza proteinelor la procariote are loc la ribozomi. Celulele procariote au puține organite.
Ipotezele originii organelelor celulelor eucariote.
Celulele procariote au apărut pe Pământ mai devreme decât celulele eucariote.
1) ipoteza simbiotică explică mecanismul de apariție a unor organite ale celulei eucariote – mitocondriile și plastidele fotosintetice.
2) Ipoteza de invaginație- afirmă că originea celulei eucariote provine din faptul că forma ancestrală a fost o procariotă aerobă. Organelele din el au apărut ca urmare a invaginării și detașării unor părți ale cochiliei, urmată de specializarea funcțională în nucleu, mitocondrii, cloroplaste ale altor organite.
§ 18. Membrane celulare. Transportul substanțelor prin membrane. Aparatul de suprafață al celulei, funcțiile sale.
Membrane celulare.
Membrane biologice- acestea sunt structuri subțiri adiacente de dimensiuni moleculare situate pe suprafața celulelor și a părților subcelulare, precum și a tubilor și veziculelor care pătrund în protoplasmă. Funcția membranelor biologice este de a regla transportul ionilor, zaharurilor, aminoacizilor și a altor produse metabolice.
Baza oricărei membrane este un strat dublu de fosfolipide.
Cu toate acestea, stratul bilipid nu este o membrană gata făcută, ci doar baza sa. Proteine numite proteine membranare. Proteinele membranare sunt cele care determină multe dintre proprietățile membranelor. Carbohidrații fac, de asemenea, parte din membrane și formează complexe cu proteine sau lipide. Membrana constă dintr-un strat bilipid în care moleculele de proteine plutesc (sau sunt fixate), formând în ea un fel de mozaic.
Structura membranei corespunde funcțiilor sale: transport, barieră și receptor.
1) Funcția de barieră. Membrana este o barieră care împiedică intrarea diferitelor substanțe chimice și a altor agenți în celule.
2) Funcțiile receptorului. Suprafața membranei are un set mare de receptori care fac posibile reacții specifice cu diverși agenți.
3) Funcția de transport. Transportul ionilor și al substanțelor are loc prin membrană.
Prin acoperirea celulei și separând-o de mediu, membranele biologice asigură integritatea celulelor și organelelor. Menține o distribuție neuniformă a ionilor de potasiu, sodiu, clor și alți ioni între protoplasmă și mediu.
O membrană deosebit de importantă în celulă este plasmalema- membrana de suprafata. Îndeplinește funcții de barieră, transport, receptor, semnalizare.
Transportul substanțelor prin membrane.
Există două procese active: exocitoză și endocitoză.
Substanțele sunt îndepărtate din celulă prin exocitoză- fuziunea veziculelor intracelulare cu membrana plasmatică. Substanțele pot intra în celulă prin endocitoza.În timpul procesului de endocitoză, membrana plasmatică formează concavități și crește, care apoi se desprind și se transformă în vezicule sau vacuole.
Există două tipuri de endocitoză:
- Pinocitoza- absorbția substanțelor lichide și dizolvate folosind bule mici;
- Fagocitoză- absorbția particulelor mari, cum ar fi microorganismele sau resturile celulare.
În cazul fagocitozei se formează bule mari, care se numesc vacuole.
Moleculele trec prin membrane prin procesele: difuzie simplă, difuzie facilitată, transport activ.
Difuziune simplă- Acesta este un exemplu de transport pasiv, trecerea dintr-o zonă cu o concentrație mai mare de molecule într-o zonă cu o concentrație mai mică. Prin difuzie simplă, substanțele nepolare (hidrofobe) solubile în lipide și molecule mici neîncărcate (de exemplu, apă) pătrund în celulă. Cu toate acestea, majoritatea substanțelor sunt transportate prin membrană folosind proteine de transport încorporate în aceasta. Există două forme de adresare: difuzie facilitată și transport activ.
Difuzare facilitată este determinată de un gradient de concentrație, iar moleculele se mișcă în funcție de acest gradient. Cu toate acestea, molecula este încărcată, transportul ei este afectat atât de gradientul de concentrație, cât și de potențialul membranei.
Transport activ este transportul substanțelor dizolvate împotriva unui gradient de concentrație folosind energia ATP. Energia este necesară deoarece materia trebuie să se miște, contrar tendinței sale naturale de a se mișca prin difuzie, în direcția opusă. Un exemplu este pompa de sodiu-potasiu. Conform legilor difuziei, ionii de Na se deplasează în mod constant în celulă, iar ionii K + ies din celulă. Încălcarea concentrației necesare a acestor ioni duce la moartea celulelor.
Aparatul de suprafață al celulei.
O varietate de celule procariote și eucariote constă din părți: aparat de suprafață, citoplasmă, aparat nuclear.
Aparatură de suprafață celulele îndeplinesc trei funcții care sunt universale pentru toate tipurile de celule: barieră, transport, receptor. De asemenea, poate îndeplini o serie de funcții specifice (de exemplu, funcția de turgență mecanică a peretelui celular din celulele vegetale). Aparatul de suprafață al celulelor este format din sisteme: membrana plasmatică, complexul supramembranar și aparatul musculo-scheletic submembranar (adică submembrană).
membrană plasmatică, sau plasmalema, este sistemul principal al aparatului de suprafață, universal pentru toate celulele. Sub acesta se află un sistem submembranar, care este implicat în transportul și recepția transmembranară și face parte din citoplasmă.
Structura supramembrana Aparatul de suprafață interacționează între celule și mediul extern sau cu alte celule. În celulele animale, complexul supramembranar sau glicocalix, joacă un rol important în funcția de receptor a celulelor. Glicocalixul este format din carbohidrați și este relativ subțire și elastic.
Aparține structurilor supramembranare derivate perete celular. Trebuie să fie produs de celulele plantelor, ciupercilor și bacteriilor. Peretele celular al plantelor conține celuloză, ciuperci - chitină, bacterii - mureină. Este destul de rigid și nu se micșorează. Apa, sărurile și moleculele multor substanțe organice trec prin peretele celular. Fenomenul de plasmoliză și deplasmoliză în celulele vegetale.
Plasmoliza- aceasta este separarea citoplasmei de membrană atunci când celula este scufundată în hipertonă, adică. concentrat din exterior, soluție. Dacă celulele animale sunt scufundate într-o soluție hipertonă, acestea se micșorează. Uneori celulele plasmolizate rămân în viață. Dacă astfel de celule sunt scufundate în apă în care concentrația de săruri este mai mică decât în celulă, are loc deplasmoliza.
Deplasmoliza- aceasta este revenirea citoplasmei celulelor vegetale dintr-o stare de plasmoliza la starea initiala.
Universitatea Tehnică de Stat din Murmansk
Departamentul de Biologie
Raport pe subiect:
„Metode de cercetare în citologie”
Efectuat:
student anul 1
Facultatea de Tehnologie
Departamentele Biologie
Serebryakova Lada Vyacheslavovna
Verificat:
Murmansk2001
Plan:
1.Ce studiază citologia?
2.Ideea că organismele constau din celule.
3. Metode de cercetare utilizate în citologie.
4. Fracționarea celulară.
5. Autoradiografie.
6. Determinarea duratei unor etape ale ciclului celular prin autoradiografie.
Citologia este știința celulelor. A apărut din alte științe biologice acum aproape 100 de ani. Pentru prima dată, informații generalizate despre structura celulelor au fost colectate într-o carte a lui J.-B. Carnoy's Biology of the Cell, publicată în 1884. Citologia modernă studiază structura celulelor, funcționarea lor ca sisteme vii elementare: funcțiile componentelor celulare individuale, procesele de reproducere celulară, repararea lor, adaptarea la condițiile de mediu și multe alte procese sunt studiate, permițând să judece proprietățile și funcțiile. comune tuturor celulelor. Citologia examinează, de asemenea, caracteristicile structurale ale celulelor specializate. Cu alte cuvinte, citologia modernă este fiziologia celulei. Citologia este strâns legată de realizările științifice și metodologice ale biochimiei, biofizicii, biologiei moleculare și geneticii. Aceasta a servit drept bază pentru un studiu aprofundat al celulei din punctul de vedere al acestor științe și apariția unei anumite științe sintetice a celulei - biologia celulară, sau biologia celulară. În prezent, termenii citologie și biologie celulară coincid, deoarece subiectul lor de studiu este celula cu propriile modele de organizare și funcționare. Disciplina „Biologie celulară” se referă la secțiunile fundamentale ale biologiei, deoarece studiază și descrie singura unitate a întregii vieți de pe Pământ – celula.
Un studiu lung și atent al celulei ca atare a condus la formularea unei importante generalizări teoretice de semnificație biologică generală și anume apariția teoriei celulare. În secolul al XVII-lea Robert Hooke, un fizician și biolog cu mare ingeniozitate, a creat un microscop. Examinând o secțiune subțire de plută la microscop, Hooke a descoperit că era format din celule minuscule goale separate de pereți subțiri, care, după cum știm acum, constau din celuloză. . El a numit aceste celule mici celule. Mai târziu, când alți biologi au început să examineze țesuturile vegetale la microscop, s-a dovedit că celulele mici descoperite de Hooke într-un dop mort, ofilit erau prezente și în țesuturile vegetale vii, dar în ele nu erau goale, ci fiecare conținea câte un corp gelatinos mic. După ce țesuturile animale au fost supuse examinării microscopice, s-a constatat că ele constau și din corpuri gelatinoase mici, dar că aceste corpuri sunt doar rareori separate între ele prin pereți.Ca urmare a tuturor acestor studii, în 1939, Schleiden și Schwann în mod independent a formulat teoria celulară, care afirmă că celulele sunt unitățile elementare din care sunt construite în cele din urmă toate plantele și toate animalele. De ceva vreme, sensul dublu al cuvântului celulă a provocat încă unele neînțelegeri, dar apoi s-a stabilit ferm în aceste mici corpuri asemănătoare jeleuului.
Înțelegerea modernă a celulelor este strâns legată de progresele tehnice și de îmbunătățirea metodelor de cercetare. Pe lângă microscopia cu lumină convențională, care nu și-a pierdut rolul, microscopia de polarizare, ultravioletă, fluorescență și contrast de fază au câștigat o importanță deosebită în ultimele decenii. Printre acestea, microscopia electronică ocupă un loc special, a cărei rezoluție a făcut posibilă pătrunderea și studierea structurii submicroscopice și moleculare a celulei. Metodele moderne de cercetare au făcut posibilă dezvăluirea unei imagini detaliate a organizării celulare.
Fiecare celulă este formată dintr-un nucleu și citoplasmă, separate între ele și de mediul extern prin membrane. Componentele citoplasmei sunt: membrana, hialoplasma, reticulul endoplasmatic si ribozomii, aparatul Golgi, lizozomii, mitocondriile, incluziunile, centrul celular, organitele specializate.
O parte a unui organism care îndeplinește o funcție specială se numește organ. Orice organ - plămân, ficat, rinichi, de exemplu - fiecare are propria sa structură specială, datorită căreia joacă un anumit rol în organism. În același mod, există structuri speciale în citoplasmă, a căror structură particulară le permite să îndeplinească anumite funcții necesare pentru metabolismul celulei; Aceste structuri se numesc organele („organe mici”).
Elucidarea naturii, funcției și distribuției organelelor citoplasmatice a devenit posibilă numai după dezvoltarea metodelor de biologie celulară modernă. Cele mai utile în acest sens au fost: 1) microscopia electronică; 2) fracționarea celulară, cu ajutorul căreia biochimiștii pot izola fracții relativ pure de celule care conțin anumite organite și, astfel, pot studia reacțiile metabolice individuale de interes pentru acestea; 3) autoradiografia, care a făcut posibilă studierea directă a reacțiilor metabolice individuale care apar în organele.
Metoda prin care organelele sunt izolate din celule se numește fracționare. Această metodă s-a dovedit a fi foarte fructuoasă, dând biochimiștilor posibilitatea de a izola diferite organele celulare într-o formă relativ pură. În plus, permite determinarea compoziției chimice a organitelor și a enzimelor pe care le conțin și, pe baza datelor obținute, să tragă concluzii despre funcțiile lor în celulă. Ca o primă etapă, celulele sunt distruse prin omogenizare într-un mediu adecvat, ceea ce asigură siguranța organelelor și împiedică agregarea lor.De foarte multe ori se folosește o soluție de zaharoză pentru aceasta. Deși mitocondriile și multe alte organite celulare rămân intacte, structurile membranare, cum ar fi reticulul endoplasmatic și membrana plasmatică, se dezintegrează în fragmente. Cu toate acestea, fragmentele de membrană rezultate se închid adesea pe ele însele, rezultând vezicule rotunde de diferite dimensiuni.
În etapa următoare, omogenatul celular este supus unei serii de centrifugări, a căror viteză și durata crește de fiecare dată; acest proces se numește centrifugare diferențială. Pe fundul tuburilor de centrifugare sunt depuse diferite organele celulare la diferite viteze de centrifugare, care depinde de mărimea, densitatea și forma organelelor. Precipitatul rezultat poate fi colectat și examinat. Structurile mai mari, mai dense, cum ar fi nucleele, sunt cele mai rapide de așezare, în timp ce structurile mai mici și mai puțin dense, cum ar fi veziculele reticulului endoplasmatic, necesită rate mai mari pentru cel mai lung timp pentru a se sedimenta. Prin urmare, la viteze mici de centrifugare, nucleele sunt sedimentate, în timp ce alte organite celulare rămân în suspensie. La viteze mai mari, mitocondriile și lizozomii precipită, iar cu centrifugare prelungită și viteze foarte mari, chiar și particule mici precum ribozomii precipită. Precipitatele pot fi examinate folosind un microscop electronic pentru a determina puritatea fracțiilor rezultate. Toate fracțiile sunt într-o oarecare măsură contaminate cu alte organite. Dacă, totuși, este posibil să se obțină o puritate suficientă a fracțiilor, acestea sunt apoi supuse analizei biochimice pentru a determina compoziția chimică și activitatea enzimatică a organitelor izolate.
Relativ recent, a fost creată o altă metodă de fracţionare a celulelor - centrifugarea cu gradient de densitate; În acest caz, centrifugarea se efectuează într-o eprubetă în care soluțiile de zaharoză cu o concentrație tot mai mare și, prin urmare, cu o densitate în creștere, sunt mai întâi stratificate una peste alta. În timpul centrifugării, organelele conținute în omogenat sunt amplasate într-un tub de centrifugă la nivelurile la care se află soluțiile de zaharoză, corespunzătoare acestora ca densitate. Această metodă oferă biochimiștilor posibilitatea de a separa organele de aceeași dimensiune, dar de densități diferite (Fig. 1.).
Autoradiografia este o metodă relativ nouă, care a extins enorm capacitățile atât ale microscopiei luminoase, cât și ale microscopiei electronice. Aceasta este o metodă extrem de modernă, datorită dezvoltării fizicii nucleare, care a făcut posibilă obținerea de izotopi radioactivi ai diferitelor elemente. Autoradiografia necesită, în special, izotopi ai acelor elemente care sunt utilizate de celulă sau care se pot lega de substanțe utilizate de celulă și care pot fi administrate animalelor sau adăugate la culturi în cantități care nu perturbă metabolismul celular normal. Deoarece un izotop radioactiv (sau substanța marcată cu acesta) participă la reacțiile biochimice în același mod ca omologul său neradioactiv și, în același timp, emite radiații, calea izotopilor în organism poate fi urmărită folosind diferite metode de detectare. radioactivitate. O modalitate de a detecta radioactivitatea se bazează pe capacitatea sa de a acționa asupra filmului fotografic precum lumina; dar radiația radioactivă pătrunde în hârtia neagră folosită pentru a proteja pelicula de lumină și are asupra peliculei același efect ca și lumina.
Pentru a detecta radiațiile emise de izotopii radioactivi pe preparatele destinate studiului cu ajutorul microscoapelor cu lumină sau electronică, preparatele sunt acoperite într-o cameră întunecată cu o emulsie fotografică specială, apoi lăsate în întuneric pentru o perioadă de timp. Apoi preparatele sunt dezvoltate (tot la întuneric) și fixate. Zonele medicamentului care conțin izotopi radioactivi afectează emulsia de bază, în care „boabe” întunecate apar sub influența radiațiilor emise. Astfel, se obțin radioautografe (în greacă. radio– radiază, autoturisme– el însuși și grapho- scrie).
La început, histologii aveau doar câțiva izotopi radioactivi; de exemplu, multe studii de autoradiografie timpurie au folosit fosfor radioactiv, iar mai târziu au fost folosiți mai mulți dintre acești izotopi; Izotopul radioactiv al hidrogenului, tritiul, a găsit o utilizare deosebit de răspândită.
Autoradiografia a fost și este încă folosită pe scară largă pentru a studia unde și cum apar anumite reacții biochimice în organism.
Compușii chimici marcați cu izotopi radioactivi care sunt utilizați pentru studiul proceselor biologice se numesc precursori.Precursorii sunt de obicei substanțe asemănătoare celor pe care organismul le obține din alimente; ele servesc ca blocuri de construcție pentru construcția țesuturilor și sunt încorporate în componente complexe ale celulelor și țesuturilor în același mod în care blocurile de construcție neetichetate sunt încorporate în ele. Componenta de țesut în care este încorporat precursorul marcat și care emite radiații se numește produs.
Celulele crescute în cultură, deși aparțin aceluiași tip, se vor afla în stadii diferite ale ciclului celular la un moment dat, cu excepția cazului în care sunt luate măsuri speciale pentru a-și sincroniza ciclurile. Cu toate acestea, prin introducerea de tritiu-timidină în celule și, ulterior, realizarea de autoradiografii, este posibilă determinarea duratei diferitelor etape ale ciclului. Momentul de apariție a unei etape - mitoza - poate fi determinat fără timidină marcată. Pentru a face acest lucru, o probă de celule din cultură este ținută sub observație într-un microscop cu contrast de fază, ceea ce face posibilă monitorizarea directă a progresului mitozei și determinarea momentului acesteia. Durata mitozei este de obicei de 1 oră, deși în unele tipuri de celule durează până la 1,5 ore.
Definiţia durationG2-perioada.
Pentru a determina durata perioadei G 2, o metodă cunoscută ca etichete puls: Timidina marcată este adăugată la cultura celulară și, după un timp scurt, mediul de cultură este înlocuit cu unul proaspăt pentru a preveni absorbția ulterioară a timidinei marcate de către celule. În acest caz, eticheta este inclusă numai în acele celule care, pe parcursul unei scurte ședințe într-un mediu cu tritiu-timidină, au fost în perioada S a ciclului celular. Proporția acestor celule este mică și doar o mică parte a celulelor va primi eticheta. În plus, toate celulele care includ eticheta vor fi în interfază - de la celulele care abia au intrat în perioada S până la cele care aproape au terminat-o în timpul expunerii la tritiu-timidină. În proba prelevată imediat după îndepărtarea timidinei marcate, eticheta este conținută numai în nucleele de interfază aparținând celulelor care au fost în perioada S în perioada de expunere la etichetă; acele celule care au fost în stare de mitoză în această perioadă rămân neetichetate.
Dacă apoi continuați să prelevați probe din cultură la anumite intervale și să pregătiți o autorradiografie pentru fiecare probă succesivă, atunci va veni un moment când eticheta începe să apară în mitoticăd-cromozomi. Etichetele vor fi incluse în toate acele celule care au fost în perioada S în timpul prezenței tritiu-timidinei în mediu, iar printre aceste celule se vor număra cele care tocmai au intrat în perioada S și cele care aproape au terminat-o. Este destul de evident că acestea din urmă vor fi primele dintre celulele marcate care vor suferi mitoză și, prin urmare, eticheta va fi detectată în cromozomii lor mitotici. Astfel, intervalul dintre 1) momentul în care timidina marcată a fost îndepărtată din cultură și 2) momentul apariției cromozomilor mitotici marcați va corespunde cu durata perioadei G 2 a ciclului celular.
Definiţia durationS-perioadă.
Deoarece celulele care se află la sfârșitul perioadei S în momentul introducerii etichetei în mediu vor fi primele care intră în mitoză, atunci, prin urmare, în acele celule în care perioada S începe imediat înainte de eticheta este eliminată, cromozomii mitotici marcați vor apărea ultimii. Prin urmare, dacă am putea determina intervalul dintre momentul intrării în mitoză a celulelor marcate primele și celulele marcate ultimele, am stabili durata perioadei S. Cu toate acestea, deși momentul în care apar pentru prima dată cromozomii mitotici marcați este ușor de determinat, momentul la care ultimele celule marcate intră în mitoză nu poate fi determinat (acest lucru este îngreunat de numărul foarte mare de celule în diviziune marcate din ultimele probe). Prin urmare, durata perioadei S trebuie determinată într-un mod diferit.
Când se examinează autoradiografiile probelor succesive de celule prelevate la intervale egale de timp, se descoperă că proporția de celule care poartă marcajul în cromozomii lor mitotici crește treptat până când literalmente toate celulele care se divid sunt etichetate. Cu toate acestea, pe măsură ce celulele termină mitoza una câte una, ele devin celule de interfază marcate. Primele care completează mitoza sunt cele ale celulelor marcate care au intrat primele în ea; și, în consecință, dintre celulele cu cromozomi mitotici marcați, ultimele care au terminat mitoza sunt cele care au intrat în ea mai târziu decât toate. Deoarece durata mitozei este întotdeauna aceeași, atunci, așadar, dacă am putea determina intervalul dintre: 1) momentul sfârșitului mitozei în celulele care au activat mai întâi marcajul și 2) momentul sfârșitului mitoza în celulele care au activat marcajul ultima, am stabili durata perioadei S. Durata S- Perioada poate fi stabilită cu ușurință prin determinarea intervalului dintre: 1) momentul în care 50% din celulele mitotice din cultură sunt nemarcate și 2) momentul după care cultura nu mai conține 50% din celulele marcate.
Determinarea timpului de generare (durata totală a întregului ciclu celular).
Continuând să prelevați probe de celule din cultură, puteți constata că la un moment dat figurile mitotice marcate dispar complet și apoi apar din nou. Astfel de celule de divizare sunt celule fiice derivate din acele celule mamă care au activat eticheta în timp ce se aflau în perioada S în momentul expunerii la tritiu-timidină. Aceste celule mamă au intrat în perioada S, s-au divizat și apoi au trecut printr-o a doua interfază și o a doua diviziune, adică au trecut printr-un ciclu complet și o parte din următorul. Timpul necesar pentru a finaliza un ciclu celular complet se numește timp generaţie. Ea corespunde intervalului dintre două vârfuri succesive de includere a etichetei și de obicei corespunde intervalului dintre acele puncte de curbe ascendente succesive în care 50% din figurile mitotice conțin eticheta.
Literatură.
A. Ham, D. Cormack „Histologie”, volumul 1 Moscova „MIR” 1982;
M.G. Abramov „Citologie clinică” Moscova „MEDICINA” 1974;
Y.S.Chentsov „Citologie generală”
Universitatea Tehnică de Stat din Murmansk
Departamentul de Biologie
Raport pe subiect:
„Metode de cercetare în citologie”
Efectuat:
student anul 1
Facultatea de Tehnologie
Departamentele Biologie
Serebryakova Lada Vyacheslavovna
Verificat:
Murmansk 2001
Plan:
1. Ce studiază citologia?
2. Ideea că organismele sunt făcute din celule.
3. Metode de cercetare utilizate în citologie.
4. Fracționarea celulară.
5. Autoradiografie.
6. Determinarea duratei unor etape ale ciclului celular prin autoradiografie.
Citologia este știința celulelor. A apărut din alte științe biologice acum aproape 100 de ani. Pentru prima dată, informații generalizate despre structura celulelor au fost colectate într-o carte a lui J.-B. Carnoy's Biology of the Cell, publicată în 1884. Citologia modernă studiază structura celulelor, funcționarea lor ca sisteme vii elementare: funcțiile componentelor celulare individuale, procesele de reproducere celulară, repararea lor, adaptarea la condițiile de mediu și multe alte procese sunt studiate, permițând să judece proprietățile și funcțiile. comune tuturor celulelor. Citologia examinează, de asemenea, caracteristicile structurale ale celulelor specializate. Cu alte cuvinte, citologia modernă este fiziologia celulei. Citologia este strâns asociată cu realizările științifice și metodologice ale biochimiei, biofizicii, biologiei moleculare și geneticii. Aceasta a servit drept bază pentru un studiu aprofundat al celulei din punctul de vedere al acestor științe și apariția unei anumite științe sintetice despre celula - biologia celulară, sau biologia celulară. În prezent, termenii citologie și biologie celulară coincid, deoarece subiectul lor de studiu este celula cu propriile modele de organizare și funcționare. Disciplina „Biologie celulară” se referă la secțiunile fundamentale ale biologiei, deoarece studiază și descrie singura unitate a întregii vieți de pe Pământ – celula.
Un studiu îndelungat și atent al celulei ca atare a condus la formularea unei importante generalizări teoretice care are semnificație biologică generală și anume apariția teoriei celulare. ÎNXVIIV. Robert Hooke, un fizician și biolog, remarcat prin mare ingeniozitate, a creat un microscop. Examinând o secțiune subțire de plută la microscop, Hooke a descoperit că era construită din celule goale mici separate de pereți subțiri, care, după cum știm acum, constau din celuloză. El a numit aceste celule mici celule. Mai târziu, când alți biologi au început să examineze țesuturile plantelor la microscop, s-a dovedit că celulele mici descoperite de Hooke într-un dop mort și ofilit erau prezente și în țesuturile vegetale vii, dar nu erau goale, ci fiecare conținea câte un mic gelatinos. corp. După ce țesuturile animale au fost supuse examinării microscopice, s-a constatat că acestea constau și din corpuri gelatinose mici, dar că aceste corpuri erau doar rareori separate între ele prin pereți. Ca rezultat al tuturor acestor studii, în 1939, Schleiden și Schwann au formulat independent teoria celulară, care afirmă că celulele sunt unitățile elementare din care sunt construite în cele din urmă toate plantele și toate animalele. De ceva vreme, sensul dublu al cuvântului celulă a provocat încă unele neînțelegeri, dar apoi s-a stabilit ferm în aceste mici corpuri asemănătoare jeleuului.
Înțelegerea modernă a celulei este strâns legată de progresele tehnice și de îmbunătățirea metodelor de cercetare. Pe lângă microscopia cu lumină convențională, care nu și-a pierdut rolul, microscopia de polarizare, ultravioletă, fluorescență și contrast de fază au câștigat o importanță deosebită în ultimele decenii. Printre acestea, microscopia electronică ocupă un loc special, a cărei rezoluție a făcut posibilă pătrunderea și studierea structurii submicroscopice și moleculare a celulei. Metodele moderne de cercetare au făcut posibilă dezvăluirea unei imagini detaliate a organizării celulare.
Fiecare celulă este formată dintr-un nucleu și citoplasmă, separate între ele și de mediul extern prin membrane. Componentele citoplasmei sunt: membrana, hialoplasma, reticulul endoplasmatic si ribozomii, aparatul Golgi, lizozomii, mitocondriile, incluziunile, centrul celular, organitele specializate.
O parte a unui organism care îndeplinește o funcție specială se numește organ. Orice organ - plămân, ficat, rinichi, de exemplu - fiecare are propria sa structură specială, datorită căreia joacă un anumit rol în organism. În același mod, există structuri speciale în citoplasmă, a căror structură particulară le oferă posibilitatea de a îndeplini anumite funcții necesare pentru metabolismul celulei; aceste structuri se numesc organele („organe mici”).
Elucidarea naturii, funcției și distribuției organelelor citoplasmatice a devenit posibilă numai după dezvoltarea metodelor de biologie celulară modernă. Cele mai utile în acest sens au fost: 1) microscopia electronică; 2) fracționarea celulară, cu ajutorul căreia biochimiștii pot izola fracții relativ pure de celule care conțin anumite organite și, astfel, pot studia reacțiile metabolice individuale de interes pentru acestea; 3) autoradiografia, care a făcut posibilă studierea directă a reacțiilor metabolice individuale care apar în organele.
Metoda prin care organelele sunt izolate din celule se numește fracționare. Această metodă s-a dovedit a fi foarte fructuoasă, dând biochimiștilor posibilitatea de a izola diferite organele celulare într-o formă relativ pură. De asemenea, permite determinarea compoziției chimice a organitelor și a enzimelor pe care le conțin și, pe baza datelor obținute, să tragă concluzii despre funcțiile lor în celulă. Ca o primă etapă, celulele sunt distruse prin omogenizare într-un mediu adecvat care păstrează organelele și împiedică agregarea acestora. Foarte des se folosește o soluție de zaharoză pentru aceasta. Deși mitocondriile și multe alte organite celulare rămân intacte, structurile membranare, cum ar fi reticulul endoplasmatic și membrana plasmatică, se dezintegrează în fragmente. Cu toate acestea, fragmentele de membrană rezultate se închid adesea pe ele însele, rezultând vezicule rotunde de diferite dimensiuni.
În etapa următoare, omogenatul celular este supus unei serii de centrifugări, a căror viteză și durata crește de fiecare dată; acest proces se numește centrifugare diferențială. Diferite organele celulare sunt depuse la fundul tuburilor de centrifugare la viteze diferite de centrifugare, care depinde de dimensiunea, densitatea și forma organelelor. Precipitatul rezultat poate fi colectat și examinat. Structurile mai mari și mai dense, cum ar fi nucleele, sunt cele mai rapide de așezare, în timp ce structurile mai mici și mai puțin dense, cum ar fi veziculele reticulului endoplasmatic, necesită viteze mai mari și timpi mai lungi de așezare. Prin urmare, la viteze mici de centrifugare, nucleele sunt sedimentate, în timp ce alte organite celulare rămân în suspensie. La viteze mai mari, mitocondriile și lizozomii precipită, iar cu centrifugare prelungită și viteze foarte mari, chiar și particule mici precum ribozomii precipită. Precipitatele pot fi examinate folosind un microscop electronic pentru a determina puritatea fracțiilor rezultate. Toate fracțiile sunt contaminate într-o oarecare măsură cu alte organite. Dacă, totuși, este posibil să se obțină o puritate suficientă a fracțiilor, acestea sunt apoi supuse analizei biochimice pentru a determina compoziția chimică și activitatea enzimatică a organitelor izolate.
Mai recent, a fost creată o altă metodă de fracţionare a celulelor - centrifugarea cu gradient de densitate; În acest caz, centrifugarea se efectuează într-o eprubetă în care soluțiile de zaharoză cu concentrații crescânde și, în consecință, cu densitate crescândă sunt mai întâi stratificate una peste alta. În timpul centrifugării, organelele conținute în omogenat sunt amplasate într-un tub de centrifugă la aceleași niveluri cu soluțiile de zaharoză corespunzătoare acestora ca densitate. Această metodă oferă biochimiștilor capacitatea de a separa organele de aceeași dimensiune, dar cu densități diferite (Fig. 1.).
Autoradiografia este o metodă relativ nouă, care a extins enorm capacitățile atât ale microscopiei luminoase, cât și ale microscopiei electronice. Aceasta este o metodă extrem de modernă, care își datorează originea dezvoltării fizicii nucleare, care a făcut posibilă obținerea de izotopi radioactivi ai diferitelor elemente. Autoradiografia necesită, în special, izotopi ai acelor elemente care sunt utilizate de celulă sau care se pot lega de substanțe utilizate de celulă și care pot fi administrate animalelor sau adăugate la culturi în cantități care nu perturbă metabolismul celular normal. Deoarece un izotop radioactiv (sau substanța marcată cu acesta) participă la reacții biochimice în același mod ca omologul său neradioactiv și, în același timp, emite radiații, calea izotopilor în organism poate fi urmărită folosind diferite metode de detectare a radioactivității . O modalitate de a detecta radioactivitatea se bazează pe capacitatea acesteia de a acționa ca lumina pe filmul fotografic; dar radiația radioactivă pătrunde în hârtia neagră folosită pentru a proteja pelicula de lumină și are asupra peliculei același efect ca și lumina.
Pentru ca radiația emisă de izotopii radioactivi să poată fi detectată pe preparatele destinate studiului cu ajutorul microscoapelor cu lumină sau electronică, preparatele sunt acoperite într-o cameră întunecată cu o emulsie fotografică specială și apoi lăsate pentru ceva timp în întuneric. Apoi preparatele sunt dezvoltate (tot la întuneric) și fixate. Zonele medicamentului care conțin izotopi radioactivi afectează emulsia de bază, în care „boabe” întunecate apar sub influența radiației emise. Astfel, se obțin radioautografe (din greacă.radio – radiază,autoturisme – el însuși șigrapho - scrie).
La început, histologii aveau doar câțiva izotopi radioactivi; de exemplu, multe studii de autoradiografie timpurie au folosit fosfor radioactiv. Mai târziu, au început să fie folosiți mult mai mulți dintre acești izotopi; Izotopul radioactiv al hidrogenului, tritiul, a găsit o utilizare deosebit de răspândită.
Autoradiografia a fost și este încă folosită pe scară largă pentru a studia unde și cum apar anumite reacții biochimice în organism.
Compușii chimici marcați cu izotopi radioactivi care sunt utilizați pentru studiul proceselor biologice sunt numiți precursori. Precursorii sunt de obicei substanțe similare cu cele pe care organismul le obține din alimente; ele servesc ca blocuri de construcție pentru construcția țesuturilor și sunt încorporate în componente complexe ale celulelor și țesuturilor în același mod în care blocurile de construcție neetichetate sunt încorporate în ele. Componenta de țesut în care este încorporat precursorul marcat și care emite radiații se numește produs.
Celulele crescute în cultură, deși aparțin aceluiași tip, se vor afla în stadii diferite ale ciclului celular la un moment dat, cu excepția cazului în care sunt luate măsuri speciale pentru a-și sincroniza ciclurile. Cu toate acestea, prin introducerea de tritiu-timidină în celule și apoi realizarea de autoradiografii, se poate determina durata diferitelor etape ale ciclului. Momentul de apariție a unei etape - mitoza - poate fi determinat fără timidină marcată. Pentru a face acest lucru, o probă de celule din cultură este ținută sub observație într-un microscop cu contrast de fază, ceea ce face posibilă monitorizarea directă a progresului mitozei și determinarea momentului acesteia. Durata mitozei este de obicei de 1 oră, deși în unele tipuri de celule durează până la 1,5 ore.
G 2-perioada .
Pentru a determina durata perioadei G 2, o metodă cunoscută caeticheta puls: Timidina marcată este adăugată la cultura celulară și, după un timp scurt, mediul de cultură este înlocuit cu unul proaspăt pentru a preveni absorbția ulterioară a timidinei marcate de către celule. În acest caz, eticheta este inclusă numai în acele celule care, pe parcursul unei scurte ședințe într-un mediu cu tritiu-timidină, au fost înS-perioada ciclului celular. Proporția acestor celule este mică și doar o mică parte a celulelor va primi eticheta. În plus, toate celulele care includ eticheta vor fi în interfază - de la celulele care abia au intratS-perioada, celor care aproape au terminat-o în timpul expunerii la tritiu-timidină. Într-o probă prelevată imediat după îndepărtarea timidinei marcate, eticheta este conținută numai în nucleele de interfază aparținând celulelor care au fost înS-perioadă; aceleași celule care au fost în stare de mitoză în această perioadă rămân neetichetate.
Dacă apoi continuați să prelevați probe din cultură la anumite intervale și să faceți o autoradiografie pentru fiecare probă succesivă, atunci va veni un moment când eticheta începe să apară înmitotică d -cromozomi . Etichetele vor fi incluse în toate acele celule care au fost în prezență de tritiu-timidină în mediu.S-punct, iar printre aceste celule vor fi cei care tocmai au intratS-perioada și aproape s-a încheiat. Este destul de evident că acestea din urmă vor fi primele dintre celulele marcate care vor suferi mitoză și, prin urmare, eticheta va fi detectată în cromozomii lor mitotici. Astfel, intervalul dintre 1) momentul în care timidina marcată a fost îndepărtată din cultură și 2) momentul apariției cromozomilor mitotici marcați va corespunde duratei.G2 perioade ale ciclului celular.
Determinarea duratei S -perioadă .
Deoarece celulele care se află la sfârșitul când eticheta este introdusă în mediuS-perioada va fi prima care va intra in mitoza, apoi, in consecinta, in acele celule in careS-perioada începe imediat înainte de îndepărtarea etichetei; cromozomii mitotici marcați vor apărea ultimii. Prin urmare, dacă am putea determina intervalul dintre momentul intrării în mitoză a celulelor marcate primele și a celulelor marcate ultimele, am stabili durata.S-perioadă. Cu toate acestea, deși momentul în care apar pentru prima dată cromozomii mitotici marcați este ușor de determinat, momentul la care ultimele celule marcate intră în mitoză nu poate fi determinat (acest lucru este îngreunat de numărul foarte mare de celule în diviziune marcate din ultimele probe). Prin urmare durataS-perioadele trebuie determinate într-un mod diferit.
La examinarea autoradiografiilor de probe succesive de celule prelevate la intervale regulate, se descoperă că proporția de celule care poartă marcajul în cromozomii lor mitotici crește treptat până când literalmente toate celulele care se divizează sunt etichetate. Cu toate acestea, pe măsură ce celulele termină mitoza una câte una, ele devin celule de interfază marcate. Primele care completează mitoza sunt cele ale celulelor marcate care au intrat primele în ea; și, în consecință, dintre celulele cu cromozomi mitotici marcați, ultimele care au terminat mitoza sunt cele care au intrat în ea mai târziu decât toate. Deoarece durata mitozei este întotdeauna aceeași, atunci, așadar, dacă am putea determina intervalul dintre: 1) momentul sfârșitului mitozei în celulele care au activat mai întâi marcajul și 2) momentul sfârșitului mitoza în celulele care s-au aprins ultimele pe marcaj, am stabili durataS-perioadă. DuratăS-perioada poate fi stabilită cu ușurință prin determinarea intervalului dintre: 1) momentul în care 50% din celulele mitotice din cultură poartă eticheta și 2) momentul după care cultura nu mai conține 50% din celule marcate.
Determinarea timpului de generare (durata totală a întregului ciclu celular).
Continuând să prelevați probe de celule din cultură, puteți constata că figurile mitotice marcate dispar complet la un moment dat și apoi apar din nou. Astfel de celule care se divide sunt celule fiice derivate din acele celule mamă care au activat eticheta în timp ce erau expuse la tritiu-timidinăS-perioadă. Aceste celule mamă au trecut înS-perioada, împărțit, apoi au trecut printr-o a doua interfază și o a doua diviziune, adică au trecut printr-un ciclu complet și o parte din următorul. Timpul necesar pentru a finaliza un ciclu celular complet se numește timpgeneraţie. Ea corespunde intervalului dintre două vârfuri succesive de încorporare a etichetei și de obicei corespunde segmentului dintre acele puncte de curbe ascendente succesive la care 50% din figurile mitotice conțin eticheta.
Literatură.
A. Ham, D. Cormack „Histologie”, volumul 1 Moscova „MIR” 1982;
M.G. Abramov „Citologie clinică” Moscova „MEDICINA” 1974;
Y.S.Chentsov „Citologie generală”
Conținutul articolului
CITOLOGIE,știința celulelor - unitățile structurale și funcționale ale aproape tuturor organismelor vii. Într-un organism multicelular, toate manifestările complexe ale vieții provin din activitatea coordonată a celulelor sale constitutive. Sarcina unui citolog este de a determina cum este construită o celulă vie și cum își îndeplinește funcțiile normale. Patomorfologii studiază și celulele, dar sunt interesați de modificările care apar în celule în timpul bolii sau după moarte. În ciuda faptului că oamenii de știință au acumulat cu mult timp în urmă o mulțime de date despre dezvoltarea și structura animalelor și plantelor, abia în 1839 au fost formulate conceptele de bază ale teoriei celulare și a început dezvoltarea citologiei moderne.
Celulele sunt cele mai mici unități ale vieții, așa cum demonstrează capacitatea țesuturilor de a se descompune în celule, care pot continua apoi să trăiască în „țesut” sau în cultura celulară și să se reproducă ca niște organisme minuscule. Conform teoriei celulare, toate organismele sunt formate din una sau mai multe celule. Există mai multe excepții de la această regulă. De exemplu, în corpul mucegaiurilor slime (mixomicete) și al unor viermi plati foarte mici, celulele nu sunt separate unele de altele, ci formează o structură mai mult sau mai puțin topită - așa-numita. sincitiu. Cu toate acestea, se poate considera că această structură a apărut secundar ca urmare a distrugerii secțiunilor de membrane celulare care au fost prezente la strămoșii evolutivi ai acestor organisme. Multe ciuperci cresc formând tuburi lungi sub formă de fire sau hife. Aceste hife, adesea împărțite prin partiții - septe - în segmente, pot fi considerate și celule alungite deosebite. Corpurile protiștilor și bacteriilor sunt formate dintr-o singură celulă.
Există o diferență importantă între celulele bacteriene și celulele tuturor celorlalte organisme: nucleele și organitele („organe mici”) ale celulelor bacteriene nu sunt înconjurate de membrane și, prin urmare, aceste celule sunt numite procariote („prenucleare”); toate celelalte celule sunt numite eucariote (cu „nuclee adevărate”): nucleele și organelele lor sunt închise în membrane. Acest articol acoperă numai celulele eucariote.
Deschiderea celulei.
Studiul celor mai mici structuri ale organismelor vii a devenit posibil abia după inventarea microscopului, adică. după 1600. Prima descriere și imagini ale celulelor au fost date în 1665 de botanistul englez R. Hooke: examinând secțiuni subțiri de plută uscată, a descoperit că acestea „constă din multe cutii”. Hooke a numit fiecare dintre aceste cutii o celulă („camera”). Cercetătorul italian M. Malpighi (1674), omul de știință olandez A. van Leeuwenhoek și englezul N. Grew (1682) au furnizat curând o mulțime de date care demonstrează structura celulară a plantelor. Cu toate acestea, niciunul dintre acești observatori nu și-a dat seama că substanța cu adevărat importantă era materialul gelatinos care umplea celulele (numit mai târziu protoplasmă), iar „celulele” care li s-au părut atât de importante erau pur și simplu cutii de celuloză fără viață care conțineau această substanță. Până la mijlocul secolului al XIX-lea. În lucrările unui număr de oameni de știință, începuturile unei anumite „teorii celulare” ca principiu structural general erau deja vizibile. În 1831, R. Brown a stabilit existența unui nucleu într-o celulă, dar nu a reușit să aprecieze întreaga importanță a descoperirii sale. La scurt timp după descoperirea lui Brown, mai mulți oameni de știință s-au convins că nucleul a fost scufundat în protoplasma semi-lichidă care umple celula. Inițial, unitatea de bază a structurii biologice a fost considerată a fi fibra. Cu toate acestea, deja la începutul secolului al XIX-lea. Aproape toată lumea a început să recunoască o structură numită veziculă, globulă sau celulă ca element indispensabil al țesuturilor vegetale și animale.
Crearea teoriei celulare.
Cantitatea de informații directe despre celulă și conținutul acesteia a crescut enorm după 1830, când au devenit disponibile microscoape îmbunătățite. Apoi, în 1838–1839, s-a întâmplat ceea ce se numește „tușa finală a maestrului”. Botanistul M. Schleiden și anatomistul T. Schwann au prezentat aproape simultan ideea structurii celulare. Schwann a inventat termenul de „teoria celulară” și a introdus această teorie în comunitatea științifică. Conform teoriei celulare, toate plantele și animalele constau din unități similare - celule, fiecare dintre ele având toate proprietățile unui lucru viu. Această teorie a devenit piatra de temelie a oricărei gândiri biologice moderne.
Descoperirea protoplasmei.
La început, s-a acordat nemeritat multă atenție pereților celulelor. Cu toate acestea, F. Dujardin (1835) a descris jeleul viu în organisme unicelulare și viermi, numindu-l „sarcoda” (adică „asemănător cu carnea”). Această substanță vâscoasă era, în opinia sa, înzestrată cu toate proprietățile viețuitoarelor. Schleiden a descoperit, de asemenea, o substanță cu granulație fină în celulele plantelor și a numit-o „mucilagiul vegetal” (1838). Opt ani mai târziu, G. von Mohl a folosit termenul „protoplasmă” (folosit în 1840 de J. Purkinje pentru a desemna substanța din care se formează embrionii de animale în stadiile incipiente de dezvoltare) și a înlocuit cu acesta termenul „mucus vegetal”. În 1861, M. Schultze a descoperit că sarcoda se găsește și în țesuturile animalelor superioare și că această substanță este identică atât structural, cât și funcțional cu așa-numita. protoplasmă vegetală. Pentru această „bază fizică a vieții”, așa cum a definit-o mai târziu T. Huxley, a fost adoptat termenul general de „protoplasmă”. Conceptul de protoplasmă a jucat un rol important la vremea sa; cu toate acestea, de mult timp a fost clar că protoplasma nu este omogenă nici ca compoziție chimică și nici ca structură, iar acest termen a căzut treptat din uz. În prezent, componentele principale ale unei celule sunt de obicei considerate a fi nucleul, citoplasma și organitele celulare. Combinația dintre citoplasmă și organele corespunde practic cu ceea ce au avut în vedere primii citologi când vorbeau despre protoplasmă.
Proprietățile de bază ale celulelor vii.
Studiul celulelor vii a aruncat lumină asupra funcțiilor lor vitale. S-a constatat că acestea din urmă pot fi împărțite în patru categorii: mobilitate, iritabilitate, metabolism și reproducere.
Motilitatea se manifestă sub diferite forme: 1) circulația intracelulară a conținutului celular; 2) flux, care asigură mișcarea celulelor (de exemplu, celulele sanguine); 3) bataia proceselor protoplasmatice minuscule - cilii si flageli; 4) contractilitatea, cea mai dezvoltată în celulele musculare.
Iritabilitatea este exprimată în capacitatea celulelor de a percepe un stimul și de a răspunde la acesta printr-un impuls sau o undă de excitare. Această activitate este exprimată în cel mai înalt grad în celulele nervoase.
Metabolismul include toate transformările materiei și energiei care au loc în celule.
Reproducerea este asigurată de capacitatea celulei de a se diviza și de a forma celule fiice. Capacitatea de a se reproduce este cea care permite celulelor să fie considerate cele mai mici unități ale vieții. Cu toate acestea, multe celule foarte diferențiate și-au pierdut această capacitate.
CITOLOGIA CA ŞTIINŢĂ
La sfârşitul secolului al XIX-lea. Atenția principală a citologilor a fost îndreptată către un studiu detaliat al structurii celulelor, al procesului de diviziune a acestora și al elucidarii rolului lor ca cele mai importante unități care furnizează baza fizică a eredității și a procesului de dezvoltare.
Dezvoltarea de noi metode.
La început, atunci când se studiază detaliile structurii celulare, a trebuit să se bazeze în principal pe examinarea vizuală a materialului mort, mai degrabă decât a materialului viu. Au fost necesare metode care să facă posibilă conservarea protoplasmei fără a o deteriora, realizarea unor secțiuni suficient de subțiri de țesut care au trecut prin componentele celulare și, de asemenea, colorarea secțiunilor pentru a dezvălui detaliile structurii celulare. Astfel de metode au fost create și îmbunătățite în a doua jumătate a secolului al XIX-lea. Microscopul în sine a fost, de asemenea, îmbunătățit. Progrese importante în designul său includ: un iluminator situat sub masă pentru a focaliza fasciculul de lumină; lentila apocromatica pentru corectarea imperfectiunilor de colorare care distorsioneaza imaginea; lentilă de imersie, oferind o imagine mai clară și o mărire de 1000 de ori sau mai mult.
De asemenea, s-a descoperit că coloranții bazici, cum ar fi hematoxilina, au o afinitate pentru conținutul nuclear, în timp ce coloranții acizi, cum ar fi eozina, colorează citoplasma; această observație a servit drept bază pentru dezvoltarea unei varietăți de metode de colorare cu contrast sau diferențial. Datorită acestor metode și microscoape îmbunătățite, cele mai importante informații despre structura celulei, „organele” sale specializate și diverse incluziuni nevii pe care celula însăși fie le sintetizează, fie le absoarbe din exterior și le acumulează treptat.
Legea continuității genetice.
Conceptul de continuitate genetică a celulelor a fost de o importanță fundamentală pentru dezvoltarea ulterioară a teoriei celulare. La un moment dat, Schleiden credea că celulele s-au format ca urmare a unui fel de cristalizare din fluidul celular, iar Schwann a mers și mai departe în această direcție eronată: în opinia sa, celulele au apărut dintr-un anumit fluid „blastema” situat în afara celulelor.
În primul rând, botaniștii și apoi zoologii (după ce contradicțiile din datele obținute din studiul anumitor procese patologice au fost clarificate) au recunoscut că celulele apar doar ca urmare a divizării celulelor deja existente. În 1858, R. Virchow a formulat legea continuității genetice în aforismul „Omnis cellula e cellula” („Fiecare celulă este o celulă”). Când s-a stabilit rolul nucleului în diviziunea celulară, W. Flemming (1882) a parafrazat acest aforism, proclamând: „Omnis nucleus e nucleo” („Fiecare nucleu este din nucleu”). Una dintre primele descoperiri importante în studiul nucleului a fost descoperirea în el a unor fire intens colorate numite cromatina. Studiile ulterioare au arătat că atunci când o celulă se divide, aceste fire sunt asamblate în corpuri discrete - cromozomi, că numărul de cromozomi este constant pentru fiecare specie, iar în procesul de diviziune celulară, sau mitoză, fiecare cromozom este împărțit în două, astfel încât fiecare celulă primește un număr tipic pentru o anumită specie de cromozomi. În consecință, aforismul lui Virchow poate fi extins la cromozomi (purtători de caracteristici ereditare), deoarece fiecare dintre ei provine dintr-unul preexistent.
În 1865 s-a stabilit că celula reproductivă masculină (spermatozoid sau spermatozoid) este o celulă cu drepturi depline, deși foarte specializată, iar 10 ani mai târziu O. Hertwig a trasat calea spermatozoizilor în procesul de fertilizare a ovulului. Și în cele din urmă, în 1884, E. van Beneden a arătat că atât în timpul formării spermatozoizilor, cât și a ovulului, are loc diviziunea celulară modificată (meioza), în urma căreia primesc un set de cromozomi în loc de doi. Astfel, fiecare spermatozoid matur și fiecare ovul matur conține doar jumătate din numărul de cromozomi în comparație cu restul celulelor unui anumit organism, iar în timpul fecundației, numărul normal de cromozomi este pur și simplu restaurat. Ca urmare, ovulul fecundat conține câte un set de cromozomi de la fiecare dintre părinți, care stă la baza moștenirii caracteristicilor atât pe liniile paterne, cât și pe cele materne. În plus, fertilizarea stimulează debutul fragmentării ovulelor și dezvoltarea unui nou individ.
Ideea că cromozomii își păstrează identitatea și mențin continuitatea genetică de la o generație de celule la alta s-a format în cele din urmă în 1885 (Rabel). S-a stabilit curând că cromozomii diferă calitativ unul de altul în influența lor asupra dezvoltării (T. Boveri, 1888). Au început să apară și date experimentale în favoarea ipotezei menționate anterior a lui V.Ru (1883), conform căreia chiar și părți individuale ale cromozomilor influențează dezvoltarea, structura și funcționarea organismului.
Astfel, chiar înainte de sfârșitul secolului al XIX-lea. s-a ajuns la două concluzii importante. Una a fost că ereditatea este rezultatul continuității genetice a celulelor furnizate de diviziunea celulară. Un alt lucru este că există un mecanism de transmitere a caracteristicilor ereditare, care este localizat în nucleu, sau mai precis, în cromozomi. S-a constatat că, datorită segregării longitudinale stricte a cromozomilor, celulele fiice primesc exact aceeași constituție genetică (atât calitativ, cât și cantitativ) ca și celula originală din care au provenit.
Legile eredității.
A doua etapă în dezvoltarea citologiei ca știință acoperă perioada 1900-1935. Ea a venit după ce legile de bază ale eredității, formulate de G. Mendel în 1865, au fost redescoperite în 1900, dar nu au atras atenția și au fost lăsate în uitare mult timp. Citologii, deși au continuat să studieze fiziologia celulei și a organelelor sale, cum ar fi centrozomul, mitocondriile și aparatul Golgi, și-au concentrat atenția principală asupra structurii cromozomilor și comportamentului lor. Experimentele de încrucișare efectuate în același timp au crescut rapid cantitatea de cunoștințe despre modurile de moștenire, ceea ce a dus la apariția geneticii moderne ca știință. Ca rezultat, a apărut o ramură „hibridă” a geneticii – citogenetica.
REALIZĂRI ALE CITOLOGIEI MODERNE
Tehnici noi, în special microscopia electronică, utilizarea izotopilor radioactivi și centrifugarea de mare viteză, dezvoltate după anii 1940, au făcut progrese enorme în studiul structurii celulelor. În dezvoltarea unui concept unificat al aspectelor fizico-chimice ale vieții, citologia se apropie din ce în ce mai mult de alte discipline biologice. În același timp, metodele sale clasice, bazate pe fixarea, colorarea și studierea celulelor la microscop, păstrează încă importanță practică.
Metodele citologice sunt utilizate, în special, în ameliorarea plantelor pentru a determina compoziția cromozomială a celulelor vegetale. Astfel de studii sunt de mare ajutor în planificarea încrucișărilor experimentale și evaluarea rezultatelor obținute. O analiză citologică similară este efectuată asupra celulelor umane: ne permite să identificăm unele boli ereditare asociate cu modificări ale numărului și formei cromozomilor. O astfel de analiză în combinație cu teste biochimice este utilizată, de exemplu, în amniocenteză pentru a diagnostica defectele ereditare ale fătului. EREDITATE.
Cu toate acestea, cea mai importantă aplicare a metodelor citologice în medicină este diagnosticarea neoplasmelor maligne. În celulele canceroase, în special în nucleele lor, apar modificări specifice care sunt recunoscute de patologii cu experiență.